Universität Bayreuth: Wie Zebrafische amputierte Flossen wiederherstellen

Bayreuther Forscher entdecken Mechanismen zur Neubildung von Knochengewebe
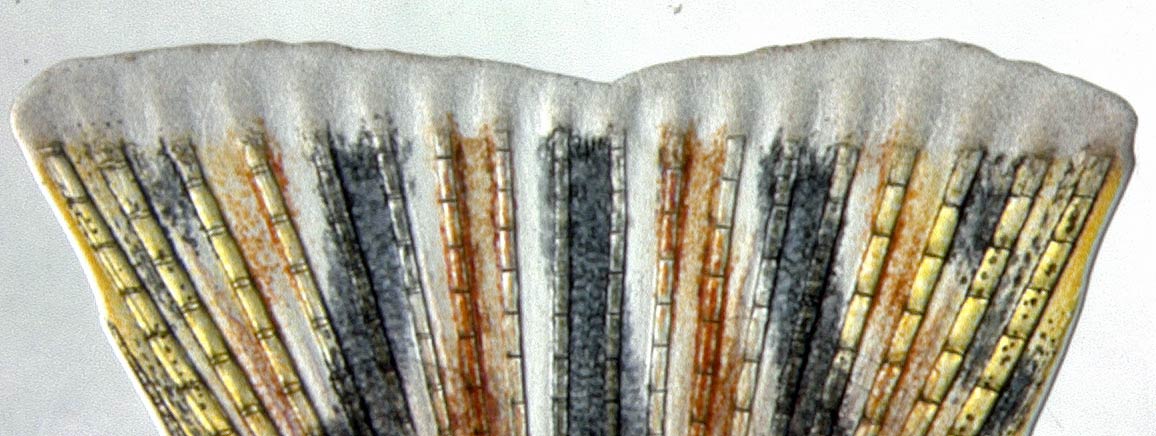
Das regenerierende Gewebe (Blastema) einer Zebrabärbling-Schwanzflosse nach einer Amputation.
Copyright: Professur für Entwicklungsbiologie, Universität Bayreuth
Im Gegensatz zum Menschen sind Fische imstande, amputierte Körperteile vollständig wiederherzustellen. Ein prominentes Beispiel ist der Zebrabärbling (Danio rerio), ein beliebter Aquarienzierfisch. Wenn seine Schwanzflossen durch Bisswunden verletzt oder im Labor amputiert werden, können sie sich innerhalb von drei Wochen vollständig regenerieren. Zebrabärblinge – auch Zebrafische genannt – bieten sich daher als Tiermodell an, um die natürliche Regeneration von Gewebe auf zellulärer und molekularer Ebene zu untersuchen.
Die Flossen der Zebrafische bestehen aus einer Flossenhaut, die durch ein Skelett aus Flossenstrahlen Stabilität bekommt; ähnlich wie ein aufgespannter Regenschirm, der durch die metallenen Kiele Festigkeit erhält. Die Flossenstrahlen werden von knochenbildenden Zellen, den Osteoblasten, gebildet. Damit eine amputierte Flosse wieder neu aufgebaut werden kann, muss in kurzer Zeit eine große Anzahl neuer Osteoblasten entstehen. Osteoblasten aus der Wundregion müssen daher die Bildung von Knochensubstanz zunächst aufgeben und sich „verjüngen“ – oder genauer gesagt: durch „De-Differenzierung“ in ein früheres Entwicklungsstadium zurückfallen. Aus spezialisierten, reifen Zellen werden teilungsfähige Knochenvorläuferzellen.
Bisher war nur wenig darüber bekannt, wie die De-Differenzierung und die erneute Knochenbildung reguliert wird. Und auch wie es dem Zebrafisch gelingt, dass das Skelett der regenerierten Flosse wieder genauso aussieht wie das der ursprünglichen Flosse, war unklar. Prof. Dr. Gerrit Begemann, Professor für Entwicklungsbiologie an der Universität Bayreuth, und seiner Doktorandin Nicola Blum ist es jetzt gelungen, wichtige Aspekte dieser Prozesse aufzuklären. In der Online-Ausgabe des renommierten Fachmagazins „Development“ haben sie dazu zwei Studien veröffentlicht. Aus den neuen Erkenntnissen können Impulse für biomedizinische Forschungsarbeiten hervorgehen, die darauf abzielen, das Gewebe verletzter Knochen oder Organe beim Menschen wiederherzustellen.
Das Dilemma der Zellen: Zwischen Vermehrung und Spezialisierung
Für das normale Wachstum der Knochen hat Retinsäure eine wichtige Funktion. Sie regen die Osteoblasten dazu an, Knochenmaterial abzuscheiden und so ihre spezielle Funktion als differenzierte Zellen zu erfüllen. Damit die Osteoblasten diese Funktion aufgeben und sich wieder zu teilungsfähigen Knochenvorläuferzellen rückentwickeln können, sind sie folglich auf eine Umgebung angewiesen, die von Retinsäure frei ist. Doch genau diese Voraussetzung ist, wenn die Schwanzflosse amputiert ist, nicht gegeben. Denn das Gewebe unter der Wunde beginnt mit der massiven Produktion von Retinsäure, um Prozesse der Zellteilung anzuregen.
Wie gelingt es den reifen Osteoblasten, diesem Dilemma zu entkommen? Die Antwort hat Nicola Blum im Labor von Prof. Gerrit Begemann entdeckt: Nach der Verletzung der Flosse produzieren die Osteoblasten vorübergehend das Enzym Cyp26b1, das Retinsäure abbaut und inaktiviert. Unter dem Schutz dieses Enzyms kann die Entwicklungsuhr zurückgedreht werden. Die teilungsfähigen Knochenvorläuferzellen wandern in einen Pool von Vorläuferzellen, das Blastema, ein. In diesem Gewebe vermehren sich die Vorläuferzellen und bilden neue Zellen für den Wiederaufbau der Flosse. In der Folge entsteht ein weiteres Dilemma, wie das Bayreuther Forschungsteam herausgefunden hat: Die Zellteilungen werden durch eine hohe Konzentration von Retinsäure unterstützt. Doch die Rückverwandlung in reife spezialisierte Zellen wird, wie schon zuvor die „De-Differenzierung“, durch Retinsäure blockiert.
Nicola Blum hat herausgefunden, wie dieses Dilemma innerhalb des Blastemas gelöst wird. In denjenigen Bereichen des Blastemas, in denen neue Osteoblasten wieder mit dem Aufbau des Skeletts beginnen, produzieren Bindegewebszellen den Retinsäurekiller Cyp26b1. Dadurch sinkt die Menge an Retinsäure, und Knochenvorläuferzellen sind in der Lage, erneut zu Osteoblasten zu reifen. Nur in demjenigen Bereich des Blastemas, der durch Zellteilung für Zellnachschub sorgt, bleibt die Konzentration an Retinsäure hoch. „So ist innerhalb des Blastemas für ein elegantes Gleichgewicht zwischen unterschiedlichen Regionen gesorgt, auf die sich die beiden Prozesse der Vermehrung und der Re-Differenzierung von Zellen verteilen“, erklärt Begemann.
Ein „Navigationssystem“ für den Wiederaufbau des Skeletts
Wie wird die Form des Knochenskeletts wiederhergestellt, sobald sich ausreichend viele Osteoblasten neu gebildet haben? Für den korrekten Wiederaufbau der Knochen ist es erforderlich, dass sich die frisch regenerierten Osteoblasten an den richtigen Stellen anlagern, nämlich exakt in der Verlängerung der noch vorhandenen Knochenstrahlen. Genau hier müssen sie neues Knochenmaterial produzieren. Der Mechanismus, der dieses Verhalten der Osteoblasten gewährleistet, war bisher unbekannt. In einer weiteren, ebenfalls in „Development“ veröffentlichten Studie konnte er von Nicola Blum aufgeklärt werden.
Damit die Osteoblasten „wissen“, wo sie sich anlagern müssen, wird in der Flossenoberhaut – auch Epidermis genannt – das Signalprotein Sonic Hedgehog produziert und ausgesendet. Die Zellen der Epidermis können dieses Protein allerdings nur dann herstellen, wenn sie frei sind von Retinsäure. Dies wird genau dort, wo Flossenstrahlen entstehen, durch das Enzym Cyp26a1 sichergestellt, das mit Cyp26b1 verwandt ist. Welche entscheidende Lotsenfunktion Sonic Hedgehog für die Osteoblasten übernimmt, konnte die Bayreuther Doktorandin zeigen, indem sie die Konzentration von Retinsäure derart manipulierte, dass das Signalprotein vom gesamten regenerierenden Gewebe ausgesendet wurde. Die Folgen waren dramatisch: Statt sich gezielt nur an der Verlängerung bestehender Flossenstrahlen anzulagern, wanderten die Osteoblasten auch in Bereiche zwischen den Strahlen ein, die normalerweise nur aus elastischer Flossenhaut besteht. Es bildeten sich Knochenzellen an völlig verkehrten Stellen, so dass im Ergebnis auch der Wiederaufbau der Skelettstruktur scheiterte.
Zugleich stellte sich in den Experimenten heraus, dass Osteoblasten ihrerseits Lotsenfunktionen für andere Zelltypen – insbesondere für Bindegewebe- und Blutgefäßzellen – haben. Auch diese Zellen müssen sich in unmittelbarer Nähe der Flossenstrahlen ansiedeln. Sie tun dies, indem sie sich an den Osteoblasten orientieren. Wenn Knochenvorläuferzellen falsche Wege einschlagen, folgen ihnen die Bindegewebe- und Blutgefäßzellen – ein weiterer Grund, weshalb die Regeneration der Skelettstruktur fehlschlägt.
„Die Neubildung des Skeletts beruht offensichtlich auf einem ‚Navigationssystem‘, das sich aus einer Kette von Lotsenfunktionen aufbaut“, fasst Begemann die neuen Erkenntnisse zusammen. „Am Beginn steht der räumlich begrenzte Abbau der Retinsäure. Dadurch wird die Produktion eines Signalproteins ermöglicht, das die knochenbildenden Zellen genau dorthin führt, wo sie benötigt werden. Die Osteoblasten ihrerseits lotsen Bindegewebe- und Blutgefäßzellen an die richtigen Stellen und koordinieren so die natürliche Regeneration der Flosse.“
Veröffentlichungen:
Osteoblast de- and redifferentiation is controlled by a dynamic response to retinoic acid during zebrafish fin regeneration. Development 2015, Vol 142 / Issue 17; posted ahead of print August 7, 2015, doi: 10.1242/dev.120204
Retinoic acid signaling spatially restricts osteoblasts and controls ray-interray organization during zebrafish fin regeneration. Development 2015, Vol 142 / Issue 17; posted ahead of print August 7, 2015, doi: 10.1242/dev.120212
Neueste Kommentare